Активация ферментов ионами металлов
Обновлено: 20.05.2024
Не менее важную роль отводят ионам металлов в осуществлении ферментативного катализа.
Участие в электрофильном катализе
Наиболее часто эту функцию выполняют ионы металлов с переменной валентностью, имеющие свободную d-орбиталь и выступающие в качестве электрофилов. В ходе электрофильного катализа ионы металлов часто участвуют в стабилизации промежуточных соединений.
Участие в окислительно-восстановительных реакциях
Ионы металлов с переменной валентностью могут также участвовать в переносе электронов. Например, в цитохромах (гемсодержащих белках) ион железа способен присоединять и отдавать один электрон:
Благодаря этому свойству цитохромы участвуют в окислительно-восстановительных реакциях.
4. Роль металлов в регуляции активности ферментов
Иногда ионы металлов выступают в роли регуляторных молекул. Например, ионы Са 2+ служат активаторами фермента протеинкиназы С, катализирующего реакции фосфорилирования белков . Ионы Са 2+ также изменяют активность ряда кальций-кальмодулинзависимых ферментов .
Б. Коферменты
Как уже было сказано, для проявления каталитической активности большинству ферментов необходимо наличие кофермента. Исключение составляют гидролитические ферменты (например, протеазы, липазы, рибонуклеаза), выполняющие свою функцию в отсутствие кофермента.
Кофермент, локализуясь в каталитическом участке активного центра, принимает непосредственное участие в химической реакции, выступая в качестве акцептора и донора химических группировок, атомов, электронов. Кофермент может быть связан с белковой частью молекулы ковалентными и нековалентными связями. В первом случае он называется простетической группой (например, FAD, FMN, биотин, липоевая кислота). Вместе с тем известны примеры, когда кофермент присоединяется к ферменту нековалентными связями настолько прочно, что не диссоциирует от белковой молекулы, например тиаминдифосфат.
Во втором случае кофермент взаимодействует с ферментом только на время химической реакции и может рассматриваться в качестве второго субстрата. Примеры - NAD + , NADP + .
Апофермент обеспечивает специфичность действия и отвечает за выбор типа химического превращения субстрата. Один и тот же кофермент, взаимодействуя с различными апоферментами, может участвовать в разных химических превращениях субстрата. Например, пиридоксальфосфат в зависимости от того, с каким апоферментом взаимодействует, участвует в реакциях трансаминирования или декарбоксилирования аминокислот.
Химическая природа коферментов, их функции в ферментативных реакциях чрезвычайно разнообразны. Традиционно к коферментам относят производные витаминов, хотя помимо них есть значительный класс небелковых соединений, принимающих участие в проявлении каталитической функции ферментов.
К коферментам относят следующие соединения:
гемы, входящие в состав цитохромов, каталазы, пероксидазы, гуанилатциклазы, NO-синтазы и являющиеся простетической группой ферментов;
нуклеотиды - доноры и акцепторы остатка фосфорной кислоты;
убихинон, или кофермент Q, участвующий в переносе электронов и протонов в ЦПЭ;
фосфоаденозилфосфосульфат, участвующий в переносе сульфата;
S-аденозилметионин (SAM) - донор метильной группы;
глутатион, участвующий в окислительно-восстановительных реакциях.
В. Мультисубстратные реакции
Большинство ферментов катализирует реакции, в которых участвует более чем один субстрат. В случае если кофермент не является простетической группой, его также можно рассматривать как ещё один субстрат. Следовательно, участников ферментативной реакции может быть несколько: непосредственно фермент, несколько субстратов и кофермент.
В этих случаях механизм ферментативной реакции, как правило, может идти по одному из двух путей: по механизму "пинг-понг" (механизму двойного замещения) или последовательному. Рассмотрим оба механизма.
Активирование и ингибирование ферментов
Скорость ферментативной реакции, как и активность фермента, в значительной степени определяется также присутствием в среде активаторов и ингибиторов: первые повышают скорость реакции, а вторые тормозят эту реакцию. Активирующее влияние на скорость ферментативной реакции оказывают разнообразные вещества органической и неорганической природы. Так, соляная кислота активирует действие пепсина желудочного сока;
желчные кислоты повышают активность панкреатической липазы; некоторые тканевые ферменты (оксидоредуктазы, катепсины, аргиназа), растительная протеиназа и др. в значительной степени активируются соединениями, содержащими свободные SH-группы (глутатион, цистеин), а ряд ферментов – также витамином С. Особенно часто активаторами выступают ионы двухвалентных и, реже, одновалентных металлов. Получены доказательства, что около четверти всех известных ферментов для проявления полной каталитической активности нуждаются в присутствии металлов. Многие ферменты вообще не активны в отсутствие металлов. Так, при удалении цинка угольная ангидраза (карбоангидраза), катализирующая биосинтез и распад Н2СО3, практически теряет свою ферментативную активность; более того, цинк при этом не может быть заменен никаким другим металлом. Известны ферменты , действие которых активируется ионами нескольких металлов; в частности, енолаза активируется Mg 2+ , Mn 2+ , К + (табл. 4.4).
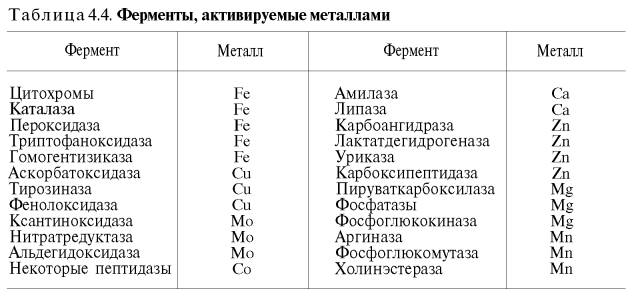
Молекулярный механизм действия металлов в энзиматическом катализе, или роль металлов в активировании ферментами. В ряде случаев ионы металлов (Со 2+ , Mg 2+ , Zn 2+ , Fe 2+ ) выполняют функции простетических групп ферментов, или служат акцепторами и донаторами электронов, или выступают в качестве электрофилов либо нуклеофилов, сохраняя реактивные группы в необходимой ориентации. В других случаях они способствуют присоединению субстрата к активному центру и образованию фермент-субстратного комплекса. Например, ионы Mg 2+ через отрицательно заряженную фосфатную группу обеспечивают присоединение монофосфатных эфиров органических веществ к активному центру фосфатаз, катализирующих гидролиз этих соединений. Иногда металл соединяется с субстратом, образуя истинный субстрат, на который действует фермент. В частности, ионы Mg 2+ активируют креатинфосфокиназу благодаря образованию истинного субстрата – магниевой соли АТФ. Наконец, имеются экспериментальные доказательства прямого участия металлов (например, ионов Са 2+ в молекуле амилазы слюны) в формировании и стабилизации активного центра и всей трехмерной структуры молекулы фермента. Следует отметить также, что металлы нередко выступают в роли аллостерических модуляторов (эффекторов; см. рис. 4.22). Взаимодействуя с аллостеричес-ким центром, подобный металл (эффектор) способствует образованию наиболее выгодной пространственной конфигурации фермента и активного фермент-субстратного комплекса.
Анионы в физиологических концентрациях обычно неэффективны или оказывают небольшое активирующее влияние на ферменты. Исключение составляют пепсин, некоторые оксидоредуктазы, активируемые анионами, а также амилаза слюны, катализирующая гидролиз крахмала, активность которой повышается при действии ионов хлора, и аденилатциклаза, которая активируется анионами галогенов.
Ингибиторы ферментов обычно принято делить на два больших класса: обратимые и необратимые. Это вещества, вызывающие частичное (обратимое) или полное торможение реакций, катализируемых ферментами. Недавно открыты антиферменты (антиэнзимы, или антизимы), представляющие собой белки (или полипептиды), действующие как ингибиторы ферментов. К подобным веществам относятся, например, ингибитор трипсина, обнаруженный в соевых бобах, и сывороточный антитрипсин. Недавно открыт в печени животных антифермент орнитинде-карбоксилазы (см. главу 12). Антизимы, вероятнее всего, образуют трудно-диссоциируемые комплексы с соответствующими ферментами, выключая их из химических реакций. Иногда ингибитор является составным компонентом предшественника фермента, например пепсина (см. главу 12), или входит в состав сложных комплексов ферментов, например в состав протеинкиназы и протеинфосфатазы, катализирующих процессы фосфо-рилирования-дефосфорилирования в живых организмах. Однако до сих пор не выяснено, являются ли подобные антиферменты истинными ингибиторами или регуляторными субъединицами, в частности, какова разница в назначении регуляторной (R) субъединицы в составе протеинкиназы и ингибиторной (I) субъединицы в составе протеинфосфатазы.
Ферменты являются белками, поэтому любые агенты, вызывающие денатурацию белка (кислоты, щелочи, соли тяжелых металлов, нагревание), приводят к необратимой инактивации фермента. Однако подобное инак-тивирование относительно неспецифично, оно не связано с механизмом действия ферментов. Гораздо большую группу составляют так называемые специфические ингибиторы, которые оказывают свое действие на какой-либо один фермент или группу родственных ферментов, вызывая обратимое или необратимое ингибирование. Исследование этих ингибиторов имеет важное значение. Во-первых, ингибиторы могут дать ценную информацию о химической природе активного центра фермента, а также о составе его функциональных групп и природе химических связей, обеспечивающих образование фермент-субстратного комплекса. Известны вещества, включая лекарственные препараты, специфически связывающие ту или иную функциональную группу в молекуле фермента, выключая ее из химической реакции. Так, йодацетат IСН2—СООН, его амид и этиловый эфир, пара-хлормеркурибензоат ClHg—С6Н4—СООН и другие реагенты сравнительно легко вступают в химическую связь с некоторыми SH-группами ферментов. Если такие группы имеют существенное значение для акта катализа, то добавление подобных ингибиторов приводит к полной потере активности фермента:
Действие ряда других ферментов (холинэстераза, трипсин и химотрип-син) сильно тормозится некоторыми фосфорорганическими соединениями, например ДФФ, вследствие блокирования ключевой гидроксильной группы серина в активном центре (см. ранее).
Во-вторых, ингибиторы нашли широкое применение в энзимологии при исследовании природы множественных форм ферментов и изоферментов, различающихся не столько электрофоретической подвижностью, сколько различной чувствительностью к одному и тому же ингибитору.
При помощи ингибиторов, выключающих отдельные стадии многоступенчатого метаболического процесса, могут быть точно установлены не только последовательность химических реакций, но и природа участвующих в этих превращениях ферментов. Этим путем, применяя йодацетат, фториды и другие специфические ингибиторы, был расшифрован глико-литический путь окислительно-восстановительных превращений глюкозы до стадии образования молочной кислоты в мышечной ткани, насчитывающий 11 стадий с участием 11 ферментов и 10 промежуточных метаболитов.
С ингибированием ферментов связан механизм действия многих токсинов и ядов на организм. Известно, что при отравлениях солями сенильной кислоты смерть наступает вследствие полного торможения и выключения дыхательных ферментов (цитохромная система) тканей, особенно клеток мозга. Токсическое влияние на организм человека и животных некоторых инсектицидов обусловлено торможением активности холинэстеразы – фермента, играющего ключевую роль в деятельности нервной системы.
Современная, так называемая рациональная, химиотерапия (направленное применение лекарственных препаратов в медицине) должна основываться на точном знании механизма действия лекарственных средств на биосинтез ферментов, на активность уже синтезированных ферментов или на регуляцию их активности в организме. Иногда для лечения некоторых болезней используют избирательно действующие ингибиторы. Так, ингибитор ряда протеиназ (трипсина, химотрипсина и калликреина) трасилол широко применяется для лечения острого панкреатита – болезни, при которой уровень трипсина и химотрипсина в крови резко возрастает. Знание избирательного ингибиторного действия некоторых природных и синтетических соединений (так называемых антиметаболитов) на ферменты может служить методологической основой для разработки эффективных методов синтеза химиотерапевтических препаратов. Этот путь открывает широкие возможности для направленного воздействия на синтез ферментов в организме и регуляции интенсивности метаболизма при патологии.
Типы ингибирования. Различают обратимое и необратимое ингибиро-вание. Если ингибитор вызывает стойкие изменения пространственной третичной структуры молекулы фермента или модификацию функциональных групп фермента, то такой тип ингибирования называется необратимым. Чаще, однако, имеет место обратимое ингибирование, поддающееся количественному изучению на основе уравнения Михаэлиса-Ментен. Обратимое ингибирование в свою очередь разделяют на конкурентное и неконкурентное в зависимости от того, удается или не удается преодолеть торможение ферментативной реакции путем увеличения концентрации субстрата.
Конкурентное ингибирование может быть вызвано веществами, имеющими структуру, похожую на структуру субстрата, но несколько отличающуюся от структуры истинного субстрата. Такое ингибирование основано на связывании ингибитора с субстратсвязывающим (активным) центром. Классическим примером подобного типа ингибирования является торможение сукцинатдегидрогеназы (СДГ) малоновой кислотой. Этот фермент катализирует окисление путем дегидрирования янтарной кислоты (сукцината) в фумаровую:
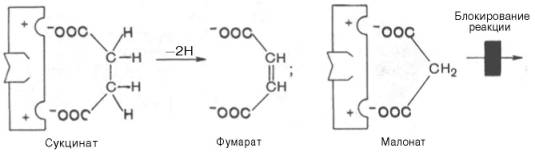
Если в среду добавить малонат (ингибитор), то в результате структурного сходства его с истинным субстратом сукцинатом (наличие двух таких же ионизированных карбоксильных групп) он будет взаимодействовать с активным центром с образованием фермент-ингибиторного комплекса, однако при этом полностью исключается перенос атома водорода от малоната. Структуры субстрата (сукцинат) и ингибитора (малонат) все же несколько различаются. Поэтому они конкурируют за связывание с активным центром, и степень торможения будет определяться соотношением концентраций малоната и сукцината, а не абсолютной концентрацией ингибитора. Таким образом, ингибитор может обратимо связываться с ферментом, образуя фермент-ингибиторный комплекс. Этот тип ингиби-рования иногда называют ингибированием по типу метаболического антагонизма (рис. 4.20).
В общей форме реакция взаимодействия ингибитора с ферментом может быть представлена следующим уравнением:
Образовавшийся комплекс, называемый фермент-ингибиторным комплексом ЕI, в отличие от фермент-субстратного комплекса ES не распадается с образованием продуктов реакции. Константу диссоциации комплекса EI, или ингибиторную константу Кi, можно, следуя теории Михаэлиса–Мен-тен, определить как отношение констант обратной и прямой реакций:
т.е. ингибиторная константа прямо пропорциональна произведению концентрации фермента и ингибитора и обратно пропорциональна концентрации комплекса EI.
Метод конкурентного торможения нашел широкое применение в медицинской практике. Известно, например, что для лечения некоторых инфекционных заболеваний, вызываемых бактериями, применяют сульфаниламидные препараты. Оказалось, что эти препараты имеют структурное сходство с парааминобензойной кислотой, которую бактериальная клетка использует для синтеза фолиевой кислоты, являющейся составной частью
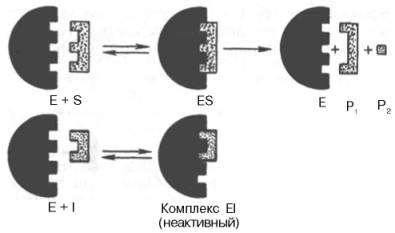
Рис. 4.20. Действие конкурентного ингибитора (схема по В.Л. Кретовичу). Е - фермент; S - субстрат; Р1 и Р2 - продукты реакции; I - ингибитор.
ферментов бактерий. Благодаря этому структурному сходству сульфаниламид блокирует действие фермента путем вытеснения парааминобензой-ной кислоты из комплекса с ферментом, синтезирующим фолиевую кислоту, что ведет к торможению роста бактерий.
Некоторые аналоги витамина В6 и фолиевой кислоты, в частности дезоксипиридоксин и аминоптерин (см. главу 7), действуют как конкурентные, так называемые коферментные, ингибиторы (или антивитамины), тормозящие многие интенсивно протекающие при патологии биологические процессы в организме. Применение подобных аналогов в медицинской практике (в частности, в дерматологии и онкологии) основано на конкурентном вытеснении коферментов из субстратсвязывающих центров ключевых ферментов обмена.
Неконкурентное ингибирование вызывается веществами, не имеющими структурного сходства с субстратами и часто связывающимися не с активным центром, а в другом месте молекулы фермента. Степень торможения во многих случаях определяется продолжительностью действия ингибитора на фермент. При данном типе ингибирования благодаря образованию стабильной ковалентной связи фермент часто подвергается полной инактивации, и тогда торможение становится необратимым. Примером необратимого ингибирования является действие йодацетата, ДФФ, а также диэтил-n-нитрофенилфосфата и солей синильной кислоты. Это действие заключается в связывании и выключении функциональных групп или ионов металлов и молекуле фермента.
Следует указать, что неконкурентное ингибирование также может быть обратимым и необратимым, поскольку отсутствует конкуренция между субстратом и ингибитором за активный центр. Примеры необратимого ингибирования приведены ранее. При обратимом неконкурентном ингибировании субстрат S и ингибитор I связываются с разными центрами, поэтому появляется возможность образования как комплекса EI, так и тройного комплекса EIS; последний может распадаться с освобождением продукта, но с меньшей скоростью, чем комплекс ES.
Этот тип неконкурентного ингибирования чаще всего наблюдается у ферментов, катализирующих превращения более одного субстрата, когда связывание ингибитора не блокирует связывание субстрата с активным центром. Ингибитор при этом соединяется как со свободным ферментом, так и с ES-комплексом.
Известно, кроме того, так называемое бесконкурентное ингиби-рование, когда ингибитор связывается с ферментом также в некаталитическом центре, однако не со свободным ферментом, а только с ES-комплексом в виде тройного комплекса.
Для выяснения вопроса о типе ингибирования пользуются уравнениями Михаэлиса-Ментен, Лайнуивера-Бэрка или другими, например уравнением Эди-Хофсти:
и соответствующими графиками в прямолинейных координатах.
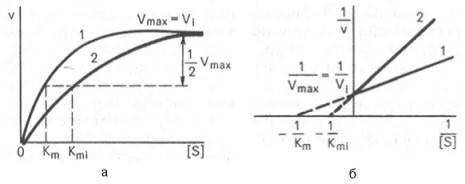
Рнс. 4.21. Графики зависимости скорости ферментативной реакции от концентрации субстрата в присутствии конкурентного ингибитора.
а - в координатах v от [ S ] ; б - в координатах 1/v от 1 / [ S ] ; Vmaxи Vi - максимальные скорости реакции; Кm и Kmi - константа Михаэлиса соответственно в отсутствие (1) и в присутствии (2) ингибитора.
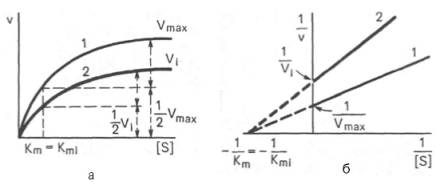
Рис. 4.22. Графики зависимости скорости ферментативной реакции от концентрации субстрата в присутствии неконкурентного ингибитора. Обозначения те же, что на рис. 4.21.
При конкурентном типе ингибирования ингибитор увеличивает значение Кm, не оказывая влияния на максимальную скорость Vmax(рис. 4.21). Это означает, что при достаточно высокой концентрации субстрата [ S ] ингибитор вытесняется молекулами субстрата из комплекса EI. При неконкурентном ингибировании (рис. 4.22) ингибитор снижает величину максимальной скорости. Если при этом величина Кm не уменьшается, то говорят о полностью неконкурентном ингибировании. Подобный тип ингибиро-вания имеет место при образовании неактивных, труднодиссоциирующих комплексов EI и(или) EIS. Часто, однако, наблюдается смешанный тип ингибирования, иногда называемый частично неконкурентным, или обратимым неконкурентным ингибированием (см. ранее), при котором снижение Vmaxсочетается с одновременным увеличением значений Кm. Это означает, что комплекс EI сохраняет частичную активность, т.е. способность к образованию промежуточного тройного комплекса EIS, в котором субстрат подвергается замедленному каталитическому превращению. В редких случаях степень торможения активности фермента может увеличиваться с повышением концентрации субстрата. Для этого типа торможения был предложен, как отмечено ранее, довольно неточный термин «бесконкурентное ингибирование». Один из механизмов такого торможения обусловлен возможностью соединения ингибитора с комплексом ES с образованием неактивного или медленно реагирующего тройного комплекса EIS.
Таким образом, при графическом анализе скоростей ферментативных реакций как функции концентраций субстрата может быть получена ценная информация не только о кинетике ферментативных реакций, но и о молекулярных механизмах ферментативного катализа.
Механизм действия активаторов
Активаторы – вещества, повышающие скорость ферментативных реакций. Различают специфические активаторы, повышающие активность одного фермента (НСl - активатор пепсиногена) и неспецифические активаторы, увеличивающие активность целого ряда ферментов (ионы Mg – активаторы гексокиназы, К, Na –АТФ-азы и других ферментов). В качестве активаторов могут служить ионы металлов, метаболиты, нуклеотиды.
- Достраивание активного центра фермента, в результате чего облегчается взаимодействие фермента с субстратом. Таким механизмом обладают в основном ионы металлов.
- Аллостерический активатор взаимодействует с аллостерическим участком (субъединицей) фермента, через его изменения опосредованно изменяет структуру активного центра и увеличивает активность фермента. Аллостерическим эффектом обладают метаболиты ферментативных реакций, АТФ.
- Аллостерический механизм может сочетаться с изменением олигомерности фермента. Под действием активатора происходит объединение нескольких субъединиц в олигомерную форму, что резко увеличивает активность фермента. Например, изоцитрат является активатором фермента ацетил-КоА карбоксилазы.
- Фосфолирирование - дефосфолирирование ферментов относится к обратимой модификации ферментов. Присоединение Н3РО4 чаще всего резко увеличивает активность фермента. Например, два неактивных димера фермента фосфорилазы соединяются с четырьмя молекулами АТФ и образуют активную тетрамерную фосфорилированную форму фермента. Фосфолирирование ферментов может сочетаться с изменением их олигомерности. В некоторых случаях фосфорилирование фермента, наоборот, снижает его активность (например, фосфорилирование фермента гликогенсинтетазы)
- Частичный протеолиз (необратимая модификация). При этом механизме от неактивной формы фермента (профермента) отщепляется фрагмент молекулы, блокирующий активный центр фермента. Например, неактивный пепсиноген под действием HCL переходит в активный пепсин.
Ингибиторы– вещества, понижающие активность фермента.
По специфичности выделяют специфичные и неспецифичные ингибиторы
По обратимости эффекта различают обратимые и необратимые ингибиторы.
По месту действия встречаются ингибиторы, действующие на активный центр и вне активного центра.
По механизму действия различают на конкурентные и неконкурентные ингибиторы.
Конкурентное ингибирование.
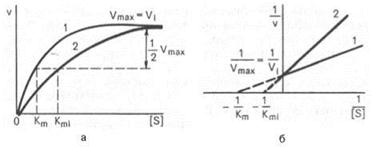
Ингибиторы этого типа имеют структуру, близкую к структуре субстрата. В силу этого ингибиторы и субстрат конкурируют за связывание активного центра фермента. Конкурентное ингибирование - это обратимое ингибирование Эффект конкурентного ингибитора может быть уменьшен путём повышения концентрации субстрата реакции
Примером конкурентного ингибирования может служить угнетение активности сукцинатдегидрогеназы, катализирующей окисление дикарбоновой янтарной кислоты, дикарбоновой малоновой кислотой, сходной по структуре с янтарной кислотой.
Принцип конкурентного ингибирования широко используется при создании лекарственных средств. Например, сульфаниламидные препараты имеют структуру, близкую к структуре парааминобензойной кислоты, необходимой для роста микроорганизмов. Сульфаниламиды блокируют ферменты микроорганизмов, необходимые для усвоения парааминобензойной кислоты. Некоторые противоопухолевые препараты являются аналогами азотистых оснований и, тем самым, ингибируют синтез нуклеиновых кислот (фторурацил).
Графически конкурентное ингибирование имеет вид:
Неконкурентное ингибирование.
Неконкурентные ингибиторы структурно не имеют схожести с субстратами реакций и поэтому не могут вытесняться при высокой концентрации субстрата. Существует несколько вариантов действия неконкурентных ингибиторов:
- Блокирование функциональной группы активного центра фермента и, вследствие этого, уменьшение активности. Например, активность SН - групп могут связывать тиоловые яды обратимо (соли металлов, ртути, свинца) и необратимо (монойодацетат). Эффект ингибирования тиоловых ингибиторов может быть уменьшен введением добавочных веществ, богатых SH группами (например, унитиол). Встречаются и используются сериновые ингибиторы, блокирующие ОН - группы активного центра ферментов. Таким эффектом обладают органические фосфофторсодержащие вещества. Эти вещества могут, в частности, ингибировать ОН - группы в ферменте ацетилхолинэстеразе, разрушающей нейромедиатор ацетилхолин.
- Блокирование ионов металлов, входящих в состав активного центра ферментов. Например, цианиды блокируют атомы железа, ЭДТА (этилендиаминтетраацетат) блокирует ионы Са, Mg.
- Аллостерический ингибитор взаимодействует с аллостерическим участком, опосредованно через него по принципу кооперативности, меняя структуру и активность каталитического участка. Графически неконкурентное ингибирование имеет вид:
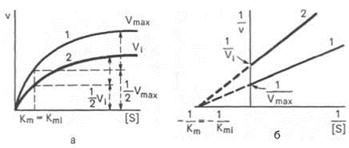
Максимальная скорость реакции при неконкурентном ингибировании не может быть достигнута путём повышения концентрации субстрата.
Молекулярный механизм действия металлов в энзиматическом катализе, или роль металлов в активировании ферментами
В ряде случаев ионы металлов (Со 2+ , Mg 2+ , Zn 2+ , Fe 2+ ) выполняют функции простетических групп ферментов, или служат акцепторами и донаторами электронов. В других случаях они способствуют присоединению субстрата к активному центру и образованию фермент-субстратного комплекса. Например, ионы Mg 2+ через отрицательно заряженную фосфатную группу обеспечивают присоединение монофосфатных эфиров органических веществ к активному центру фосфатаз, катализирующих гидролиз этих соединений. Иногда металл соединяется с субстратом, образуя истинный субстрат, на который действует фермент. В частности, ионы Mg 2+ активируют креатинфосфокиназу благодаря образованию истинного субстрата – магниевой соли АТФ. Наконец, имеются экспериментальные доказательства прямого участия металлов (например, ионов Са 2+ в молекуле амилазы слюны) в формировании и стабилизации активного центра и всей трехмерной структуры молекулы фермента.
Активаторы ферментов.Ферменты-протеинкиназы имеют четвертичную структуру, состоят их 2-х субъединиц - каталитической и регуляторной. Активатор воздействует на регуляторную, а в результате обнажается каталитическая субъединица:
Рисунок 21-Воздействие активатора
Кооперативный эффект.Характерен для ферментов, имеющих 2 и более субъединиц. Присоединение субстрата к одной из субъединиц облегчает последующие присоединения к оставшимся субъединицам. Типичный пример кооперативного эффекта - перенос кислорода молекулами гемоглобина.
Ингибиторы ферментов обычно принято делить на два больших класса: обратимые и необратимые. Это вещества, вызывающие частичное (обратимое) или полное торможение реакций, катализируемых ферментами. Недавно открыты антиферменты (антиэнзимы), представляющие собой белки (или полипептиды), действующие как ингибиторы ферментов. К подобным веществам относятся, например, ингибитор трипсина, обнаруженный в соевых бобах, и сывороточный антитрипсин. Недавно открыт в печени животных антифермент орнитиндекарбоксилазы. Антиэнзимы, вероятнее всего, образуют труднодиссоциируемые комплексы с соответствующими ферментами, выключая их из химических реакций. Иногда ингибитор является составным компонентом предшественника фермента или входит в состав сложных комплексов ферментов, например в состав протеинкиназы и протеинфосфатазы, катализирующих процессы фосфорилирования-дефосфорилирования в живых организмах.
Ферменты являются белками, поэтому любые агенты, вызывающие денатурацию белка (кислоты, щелочи, соли тяжелых металлов, нагревание), приводят к необратимой инактивации фермента. Однако подобное инак-тивирование относительно неспецифично, оно не связано с механизмом действия ферментов. Гораздо большую группу составляют так называемые специфические ингибиторы, которые оказывают свое действие на какой-либо один фермент или группу родственных ферментов, вызывая обратимое или необратимое ингибирование. Ингибиторы могут дать ценную информацию о химической природе активного центра фермента, а также о составе его функциональных групп и природе химических связей, обеспечивающих образование фермент-субстратного комплекса.
Действие ряда ферментов (холинэстераза, трипсин и химотрипсин) сильно тормозится некоторыми фосфорорганическими соединениями, например ДДТ, вследствие блокирования ключевой гидроксильной группы серина в активном центре.
При помощи ингибиторов, выключающих отдельные стадии многоступенчатого метаболического процесса, могут быть точно установлены не только последовательность химических реакций, но и природа участвующих в этих превращениях ферментов. Этим путем, применяя йодацетат, фториды и другие специфические ингибиторы, был расшифрован гликолитический путь окислительно-восстановительных превращений глюкозы до стадии образования молочной кислоты в мышечной ткани, насчитывающий 11 стадий с участием 11 ферментов и 10 промежуточных метаболитов.
Типы ингибирования.Различают обратимое и необратимое ингибирование. Если ингибитор вызывает стойкие изменения пространственной третичной структуры молекулы фермента или модификацию функциональных групп фермента, то такой тип ингибирования называется необратимым.
Чаще, однако, имеет место обратимое ингибирование, поддающееся количественному изучению на основе уравнения Михаэлиса-Ментен. Обратимое ингибирование в свою очередь разделяют на конкурентное и неконкурентное в зависимости от того, удается или не удается преодолеть торможение ферментативной реакции путем увеличения концентрации субстрата.
Конкурентное ингибирование может быть вызвано веществами, имеющими структуру, похожую на структуру субстрата, но несколько отличающуюся от структуры истинного субстрата. Такое ингибирование основано на связывании ингибитора с субстратсвязывающим (активным) центром. Классическим примером подобного типа ингибирования является торможение сукцинатдегидрогеназы (СДГ) малоновой кислотой. Этот фермент катализирует окисление путем дегидрирования янтарной кислоты (сукцината) в фумаровую:
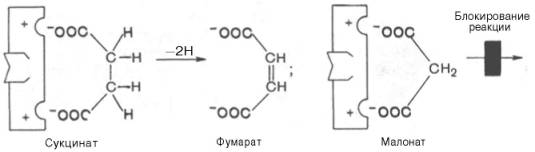
Если в среду добавить малонат (ингибитор), то в результате структурного сходства его с истинным субстратом сукцинатом (наличие двух таких же ионизированных карбоксильных групп) он будет взаимодействовать с активным центром с образованием фермент-ингибиторного комплекса, однако при этом полностью исключается перенос атома водорода от малоната. Структуры субстрата (сукцинат) и ингибитора (малонат) все же несколько различаются. Поэтому они конкурируют за связывание с активным центром, и степень торможения будет определяться соотношением концентраций малоната и сукцината, а не абсолютной концентрацией ингибитора. Таким образом, ингибитор может обратимо связываться с ферментом, образуя фермент-ингибиторный комплекс. Этот тип ингибирования иногда называют ингибированием по типу метаболического антагонизма (рис. 22).
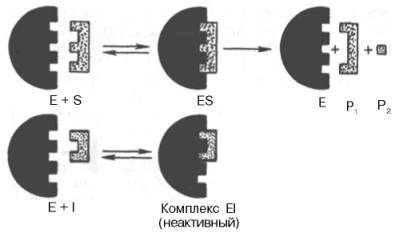
Е - фермент; S - субстрат; Р1 и Р2 - продукты реакции; I - ингибитор
Рисунок 22-Действие конкурентного ингибитора (схема по В.Л. Кретовичу)
Образовавшийся комплекс, называемый фермент-ингибиторным комплексом ЕI, в отличие от фермент-субстратного комплекса ES не распадается с образованием продуктов реакции. Константу диссоциации комплекса EI, или ингибиторную константу Кi, можно, следуя теории Михаэлиса–Ментен, определить как отношение констант обратной и прямой реакций:
т.е. ингибиторная константа прямо пропорциональна произведению концентрации фермента и ингибитора и обратно пропорциональна концентрации комплекса EI.
Метод конкурентного торможения нашел широкое применение в медицинской практике. Известно, например, что для лечения некоторых инфекционных заболеваний, вызываемых бактериями, применяют сульфаниламидные препараты. Оказалось, что эти препараты имеют структурное сходство с парааминобензойной кислотой, которую бактериальная клетка использует для синтеза фолиевой кислоты, являющейся составной частью ферментов бактерий. Благодаря этому структурному сходству сульфаниламид блокирует действие фермента путем вытеснения парааминобензойной кислоты из комплекса с ферментом, синтезирующим фолиевую кислоту, что ведет к торможению роста бактерий.
Некоторые аналоги витамина В6 и фолиевой кислоты, в частности дезоксипиридоксин, действуют как конкурентные, так называемые коферментные, ингибиторы (или антивитамины), тормозящие многие интенсивно протекающие при патологии биологические процессы в организме. Применение подобных аналогов в медицинской практике (в частности, в дерматологии и онкологии) основано на конкурентном вытеснении коферментов из субстратсвязывающих центров ключевых ферментов обмена.
Неконкурентное ингибирование вызывается веществами, не имеющими структурного сходства с субстратами и часто связывающимися не с активным центром, а в другом месте молекулы фермента. Степень торможения во многих случаях определяется продолжительностью действия ингибитора на фермент. При данном типе ингибирования благодаря образованию стабильной ковалентной связи фермент часто подвергается полной инактивации, и тогда торможение становится необратимым. Примером необратимого ингибирования является действие йодацетата, ДФФ, а также диэтил-n-нитрофенилфосфата и солей синильной кислоты. Это действие заключается в связывании и выключении функциональных групп или ионов металлов и молекуле фермента.
Известно, кроме того, так называемое бесконкурентное ингибирование, когда ингибитор связывается с ферментом также в некаталитическом центре, однако не со свободным ферментом, а только с ES-комплексом в виде тройного комплекса.
Мультимолекулярные ферментные системы.Особую группу ферментов составляют надмолекулярные (или мульти-молекулярные) ферментные комплексы, в состав которых входят не субъединицы (в каталитическом отношении однотипные протомеры), а разные ферменты, катализирующие последовательные ступени превращения какого-либо субстрата. Отличительными особенностями подобных мультиферментных комплексов являются прочность ассоциации ферментов и определенная последовательность прохождения промежуточных стадий во времени, обусловленная порядком расположения каталитически активных (различных) белков в пространстве («путь» превращения в пространстве и времени). Типичными примерами подобных мультиферментных комплексов являются пируватдегидрогеназа и α-кетоглутаратдегидрогеназа, катализирующие соответственно окислительное декарбоксилирование пировиноградной и α-кетоглутаровой кислот в животных тканях, и синтетаза высших жирных кислот. Молекулярные массы этих комплексов в зависимости от источника их происхождения варьируют от 2,3•10 6 до 10•10 6 .
Ряд таких мультиферментных комплексов, иногда называемых ферментными ансамблями, структурно связанных с какой-либо органеллой (рибосомы, митохондрии) или с биомембраной и составляет высокоорганизованные надмолекулярные системы, обеспечивающие жизненно важные функции, например, тканевое дыхание (перенос электронов от субстратов к кислороду через систему дыхательных ферментов).
Активаторы и ингибиторы ферментов
Регуляция активности ферментов может осуществляться путём взаимодействия ферментов с различными биологическими компонентами или чужеродными соединениями, которые называются регуляторами ферментов. Они могут либо ускорять, либо замедлять ферментативную реакцию.
Активаторы– это вещества,увеличивающие скорость ферментативной реакции.
Виды активаторов:
1. Вещества, влияющие на область активного центра. К ним относятся ионы металлов (Na + , K + , Fe 2+ , Co 2+ , Cu 2+ , Ca 2+ , Zn 2+ , Mg 2+ , Mn 2+ и др.). В ряде случаев ионы металлов выполняют функцию кофактора фермента. В других случаях они способствуют присоединению субстрата к активному центру фермента. Ионы металлов оказываются активаторами только в условиях дефицита их в организме.
2. Аллостерические эффекторы, которые связываются с аллостерическим (регуляторным) участком апофермента. Это связывание вызывает конформационные изменения в молекуле белка, приводящие к изменению структуры активного центра, что сказывается на связывании и превращении субстрата в активном центре. При этом активность фермента либо увеличивается (это аллостерические активаторы), либо уменьшается (это аллостерические ингибиторы). Аллостерическими эффекторами ферментов наиболее часто выступают различные метаболиты, а также гормоны, ионы металлов, нуклеозиды - АТФ, АДФ, АМФ.
3. Вещества, вызывающие модификации, не затрагивающие активный центр фермента. Возможно несколько вариантов таких модификаций:
- активация путём присоединения специфической модифицирующей группы к молекуле фермента. Пример: регуляция активности липазы.
неактивная АТФ АДФ активная О
В этом случае фосфатная группа присоединяется к гидроксильным группам аминокислот, находящихся в белковой части фермента. Отрицательно заряженные фосфатные группы могут разрывать слабые водородные и ионные связи в третичной структуре белка-фермента и влиять на конформационное состояние его активного центра. В зависимости от природы фермента фосфорилирование может его активировать или, наоборот, инактивировать. Реакции присоединения фосфатной группы катализируют ферменты протеинкиназы, а отщепления – фосфатазы. Активность этих ферментов в свою очередь находится под контролем гормональной системы.
- активация путёмперехода неактивного предшественника - профермента в активный фермент за счёт частичного протеолиза.
Некоторые ферменты синтезируются в клетке первоначально неактивными и после секреции из клетки переходят в активную форму. Неактивные предшественники называются проферменты (зимогены). Под действием активатора происходит частичный гидролиз профермента с отщеплением от него неактивного пептида, в результате чего открывается активный центр. Так происходит активация ферментов желудочно-кишечного тракта, переваривающих белки пищи. Например, фермент пепсиноген, синтезированный в клетках желудка, затем в просвете желудка под действием соляной кислоты превращается в активный пепсин путём удаления неактивного участка полипептидной цепи:
неактивный HCl активный
пепсиноген пепсин + пептид
- активатор вызывает диссоциацию субъединиц фермента, имеющего четвертичную структуру (отщепление одной из субъединиц фермента).
Ингибитораминазывают вещества, вызывающие снижение активности фермента. Следует различать инактивацию и ингибирование фермента. Сам по себе факт торможения ферментативной реакции в присутствии какого-либо вещества ещё не говорит о том, что это вещество – ингибитор. Любые денатурирующие агенты вызывают инактивацию фермента и торможение ферментативной реакции. Ингибиторы, в отличие от денатурирующих агентов, действуют в малых концентрациях и вызывают специфическое снижение ферментативной активности.
По прочности связывания с ферментом ингибиторы делятся на обратимые и необратимые. Необратимые ингибиторы прочно связываются с ферментом, тогда как комплекс фермент – обратимый ингибитор непрочен. Если сильно разбавить раствор фермента с обратимым ингибитором, то их комплекс распадается и активность фермента восстанавливается.
По механизму действия ингибиторы делятся на конкурентные и неконкурентные. Конкурентные ингибиторы имеют структурное сходство с молекулой субстрата, что позволяет им занять место субстрата в активном центре фермента:
Встраиваясь вместо субстрата в активный центр, такой ингибитор не даёт ферментативной реакции осуществиться. То есть, субстрат конкурирует с ингибитором за активный центр. С активным центром связывается то соединение, молекул которого больше. Снять конкурентное ингибирование можно, увеличив концентрацию субстрата.
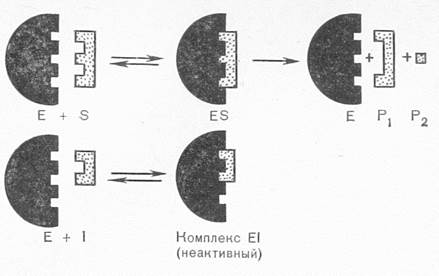
На принципе конкурентного ингибирования основано действие многих фармакологических препаратов (например, сульфаниламидных), инсектицидов, фосфорорганических боевых отравляющих веществ (зарин, зоман).
Неконкурентные ингибиторы не имеют структурного сходства с субстратами. Они или связываются с каталитическими группами активного центра фермента, или, связываясь с ферментом вне активного центра, изменяют конформацию активного центра таким образом, что это препятствует превращению субстрата. Поскольку неконкурентный ингибитор не влияет на связывание субстрата, то в отличие от конкурентного ингибирования наблюдается образование тройного комплекса:
К неконкурентным ингибиторам относятся ионы тяжёлых металлов: ртути, свинца, кадмия, мышьяка. Они блокируют SH-группы, входящие в каталитический участок фермента. Снять действие неконкурентного ингибитора избытком субстрата, как при конкурентном ингибировании, нельзя, а можно лишь веществами, связывающими ингибитор (реактиваторами). Тяжелые металлы лишь в небольших концентрациях играют роль ингибиторов, в больших концентрациях они действуют как денатурирующие агенты.
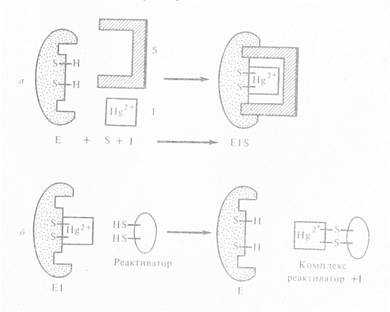
Наиболее важными неконкурентными ингибиторами являются образующиеся в живой клетке промежуточные продукты метаболизма, способные обратимо связываться с аллостерическими участками фермента – аллостерические ингибиторы. Они занимают ключевое положение в метаболизме, поскольку тонко реагируют на изменения в обмене веществ и регулируют прохождение веществ по целой системе ферментов. Например, аллостерическая регуляция проявляется в виде ингибирования конечным продуктом первого фермента цепи. Эта регуляция сходна с регуляцией по механизму обратной связи и позволяет контролировать выход конечного продукта, в случае накопления которого прекращается работа первого фермента цепи:
Е1, Е2, Е3 – ферменты; А, В, С, D - метаболиты
Читайте также: