Металл в составе хлорофилла
Обновлено: 18.05.2024
Хлорофилл – сложный эфир дикарбоновой кислоты и двух остатков спиртов (метилового, фитола).
Образование хлорофилла осуществляется в 2 фазы: первая фаза – темновая, во время которой образуется предшественник хлорофилла – протохлорофилл, а вторая – световая, при которой из протохлорофилла на свету образуется хлорофилл. Для образования хлорофилла необходимо наличие железа. При недостатке железа получаются растения, характеризующиеся бледными полосами и слабой зелёной окраской листьев. Образование хлорофилла зависит от температуры. Оптимальная температура для накопления хлорофилла 26-30°С. Как и следовало ожидать, от температуры зависит лишь образование протохлорофилла (темновая фаза). При наличии уже образовавшихся протохлорофиллов процесс зеленения (световая фаза) идёт с одинаковой скоростью независимо от температуры. На скорость образования хлорофилла оказывает влияние содержания воды. Сильное обезвоживание проростков приводит к полному прекращению образования хлорофилла. Особенно чувствительно к обезвоживанию образование протохлорофилла.
Важнейшее значение для образования хлорофилла имеют условия минерального питания. Прежде всего, необходимо достаточное количества железа. При недостатке железа даже листья взрослых растений теряют окраску.
Большое значение для обеспечения синтеза хлорофилла имеет нормальное снабжение растений азотом и магнием, так как оба эти элемента входят в состав хлорофилла. При недостатке меди хлорофилл легко разрушается. Это, по-видимому, связано с тем, что медь способствует образованию устойчивых комплексов между хлорофиллом и соответствующими белками.
Химические свойства хлорофилла.
По химическому составу хлорофилл представляет сложный эфир дикарбоновой кислоты хлорофинилла. Хлорофинилл представляет собой азотосодержащее металлоорганическое соединение, относящееся к магний-порфиринам. В центре молекуле хлорофилла расположен атом магния, который соединён с четырьмя азотами пиррольных группировок. В пиррольных группировках хлорофилла имеется система чередующихся двойных и простых связей. Это и есть хромофорная группа хлорофилла, обуславливающая его окраску.
1. Благодаря молекуле фитола молекула хлорофилла обладает липофильными свойствами.
2. Растворяется в 96%-ном спирте, ацетоне, бензоле, не растворяется в воде, фитролейном эфире.
3. Голова хлорофилла располагается на той стороне мембраны, которая обращена к водной строме.
Оптические свойства хлорофилла.
Хлорофилл поглощает солнечную энергию и направляет её на химические реакции, которые не могут протекать без энергии, получаемой извне. Раствор хлорофилла в проходящем свете имеет зелёный цвет, но при увеличении толщины слоя или концентрации хлорофилла он приобретает красный цвет.
Хлорофилл поглощает свет не сплошь, а избирательно. При пропускании из семи видимых цветов, которые постепенно переходят друг в друга. При пропускании белого света через призму и раствор хлорофилла на полученном спектре наиболее интенсивное поглощение будет в красных и сине-фиолетовых лучах. Зелёные лучи поглощаются мало, поэтому в тонком слое хлорофилл имеет в проходящем свете зелёный цвет. Однако с увеличением концентрации хлорофилла полосы поглощения расширяются и без поглощения проходит только часть крайних красных.
Спектры поглощения хлорофилла а и b очень близки.
В отражённом свете хлорофилл, кажется вишнёво-красным, так как он излучает поглощённый свет с изменением длины его волны. Это свойство хлорофилла называется флюоресценцией.
Хлорофилл
Хлорофи́лл (от греч. χλωρός , «зелёный» и φύλλον , «лист») — зелёный пигмент, окрашивающий хлоропласты растений в зелёный цвет. При его участии происходит фотосинтез. По химическому строению хлорофиллы — магниевые комплексы различных тетрапирролов. Хлорофиллы имеют порфириновое строение и близки гему.
Содержание
- 1 История открытия
- 2 В природе
- 3 Синтез
- 4 Свойства и функция при фотосинтезе
- 5 Химическая структура
- 6 Применение
История открытия
В 1817 году Жозеф Бьенеме Каванту и Пьер Жозеф Пеллетье выделили из листьев растений зелёный пигмент, который они назвали хлорофиллом. В 1900-х годах Михаил Цвет и Рихард Вильштеттер независимо обнаружили, что хлорофилл состоит из нескольких компонентов. Вильтштеттер очистил и кристаллизовал два компонента хлорофилла, названные им хлорофиллами а и b и установил брутто-формулу хлорофилла а. В 1915 году за исследования хлорофилла ему была вручена Нобелевская премия. В 1940 Ханс Фишер, получивший в 1930 Нобелевскую премию за открытие структуры гема, установил химическую структуру хлорофилла a. Его синтез был впервые осуществлен в 1960 Робертом Вудвордом, а в 1967 была окончательно установлена его стереохимическая структура.
В природе


Хлорофилл присутствует во всех фотосинтезирующих организмах — высших растениях, водорослях, сине-зелёных водорослях (цианобактериях), фотоавтотрофных простейших (протистах) и бактериях.
Некоторые высшие растения, наоборот, лишены хлорофилла (как, например, петров крест).
Синтез
Синтезирован Робертом Вудвордом в 1960 году.
Синтез включает в себя 15 реакций, которые можно разделить на 3 этапа. Исходными веществами для синтеза хлорофилла являются глицин и ацетат. На первом этапе образуется аминолевулиновая кислота. На втором этапе происходит синтез одной молекулы протопорфирина из четырёх пиррольных колец. Третий этап представляет собой образование и превращение магнийпорфиринов.
Свойства и функция при фотосинтезе
В процессе фотосинтеза молекула хлорофилла претерпевает изменения, поглощая световую энергию, которая затем используется в фотохимической реакции взаимодействия углекислого газа и воды с образованием органических веществ (как правило, углеводов):
После передачи поглощенной энергии молекула хлорофилла возвращается в исходное состояние.
Хотя максимум непрерывного спектра солнечного излучения расположен в «зелёной» области 550 нм (где находится и максимум чувствительности глаза), поглощается хлорофиллом преимущественно синий, частично — красный свет из солнечного спектра (чем и обуславливается зелёный цвет отражённого света).
Растения могут использовать и свет с теми длинами волн, которые слабо поглощаются хлорофиллом. Энергию фотонов при этом улавливают другие фотосинтетические пигменты, которые затем передают энергию хлорофиллу. Этим объясняется разнообразие окраски растений (и других фотосинтезирующих организмов) и её зависимость от спектрального состава падающего света.
Химическая структура
Хлорофиллы можно рассматривать как производные протопорфирина — порфирина с двумя карбоксильными заместителями (свободными или этерифицированными). Так, хлорофилл a имеет карбоксиметиловую группу при C10, фитоловый эфир пропионовой кислоты — при С7. Удаление магния, легко достигаемое мягкой кислотной обработкой, дает продукт, известный как феофитин. Гидролиз фитоловой эфирной связи хлорофилла приводит к образованию хлорофиллида (хлорофиллид, лишенный атома металла, известен как феофорбид a).
Все эти соединения интенсивно окрашены и сильно флуоресцируют, исключая те случаи, когда они растворены в органических растворителях в строго безводных условиях. Они имеют характерные спектры поглощения, пригодные для качественного и количественного определения состава пигментов. Для этой же цели часто используются также данные о растворимости этих соединений в соляной кислоте, в частности для определения наличия или отсутствия этерифицированных спиртов. Хлороводородное число определяется как концентрация HCl (%, масс./об.), при которой из равного объёма эфирного раствора пигмента экстрагируется 2 /3 общего количества пигмента. «Фазовый тест» — окрашивание зоны раздела фаз — проводят, подслаивая под эфирный раствор хлорофилла равный объём 30%-го раствора KOH в MeOH. В интерфазе должно образовываться окрашенное кольцо. С помощью тонкослойной хроматографии можно быстро определять хлорофиллы в сырых экстрактах.
Хлорофиллы неустойчивы на свету; они могут окисляться до алломерных хлорофиллов на воздухе в метанольном или этанольном растворе.
Хлорофиллы образуют комплексы с белками in vivo и могут быть выделены в таком виде. В составе комплексов их спектры поглощения значительно отличаются от спектров свободных хлорофиллов в органических растворителях.
Хлорофиллы можно получить в виде кристаллов. Добавление H2O или Ca 2+ к органическому растворителю способствует кристаллизации.
ХЛОРОФИЛЛЫ
ХЛОРОФИЛЛЫ (греческий chloros зеленый + phyllon лист) — пигменты растений, а также некоторых микроорганизмов, с помощью которых улавливается энергия солнечного света и осуществляется процесс фотосинтеза. Участвуя в фотосинтезе (см.), хлорофиллы играют огромную биол. роль.
Существует четыре вида хлорофиллов: a, b, c и d. Высшие растения содержат хлорофиллы a и b, бурые и диатомовые водоросли — хлорофиллы а и с, красные водоросли — хлорофилл d. Кроме того, некоторые фотосинтезирующие бактерии содержат аналоги хлорофиллов — бактериохлорофиллы. В основе молекул хлорофиллов лежит магниевый комплекс порфиринового цикла (см. Порфирины). К одному из пиррольных колец присоединен остаток многоатомного спирта фитола, благодаря чему хлорофиллы получили возможность встраиваться в липидный слой мембраны хлоропластов.
Выделение хлорофиллов в чистом виде и разделение их на два компонента (хлорофиллы а и b) впервые было осуществлено русским ботаником М. С. Цветом с помощью разработанного им метода хроматографии (см.). Им же было доказано, что в листьях растений хлорофиллы сопровождает ряд желтых спутников — каротиноидов (см.). Структурная формула хлорофиллов установлена Фишером (Н. Fischer) в 1940 году М. В. Ненцкий и его ученики доказали хим. родство гемоглобина (см.) и хлорофиллов растений. В изучении физиологической роли хлорофиллов большое значение имели исследования К. А. Тимирязева. Полный синтез хлорофиллов произвели независимо друг от друга Штрелль (М. Strell) и Вудворд (R. В. Woodword) в 1960 году.
Хлорофиллы являются главной составной частью пигментного аппарата высших растений, мхов, водорослей, фотосинтезирующих бактерий. Содержание их в растениях зависит от вида растения, обеспеченности минеральным питанием и других условий. Количество хлорофиллов в растениях колеблется от 1,7 до 5% в пересчете на сухой вес. Концентрация их на поверхности листа определяет интенсивность поглощения растением света, если уровень хлорофиллов не превышает 2 мг/дм 2 . При содержании хлорофиллов от 3 мг/дм 2 и выше коэффициент поглощения света приближается к 97 —100% и не зависит от количества пигмента.
В клетках зеленого листа хлорофиллы находятся в особых органеллах — пластидах, которые называются также хлорофилловыми зернами, или хлоропластами. Каждый хлоропласт растения Mnium medium имеет объем 4,1 X 10 -11 см 3 и содержит 1,3*10 9 молекул хлорофилла, ограничен двойной липопротеидной мембраной и заполнен белковой стромой. Чередующиеся пластинки белка и окрашенных пигментно-липидных слоев образуют включения в строме (граны). Расстояния между молекулами пигмента в тонком моно- или бимолекулярном слоях невелики; каждая из пары молекул может быть связана с ферментами типа цитохрома (см. Цитохромы), способного отдавать электрон хлорофиллу, а другая — с акцептором электрона типа ферредоксина.
Процесс фотосинтеза начинается с поглощения кванта света пигментной системой растения. Участие промежуточных систем в цепи переноса электрона показано на схеме:

где X — хлорофилл, ЦИТ — цитохромы, ФД — ферредоксин, ФЛ — флавиновые системы, hv — квант света.
Важное значение в функционирующей фотосинтетической единице имеет процесс миграции энергии между различными формами хлорофилла. Активно функционирующая фотосинтетическая единица содержит 200—400 молекул хлорофилла, которые работают как единая светоулавливающая система, поглощающая один квант света. За один цикл работы на каждые 3000 молекул хлорофилла высвобождается одна молекула кислорода. Установлено, что спектрально различные формы хлорофилла образуют лестницу энергетических уровней, по которой поглощенная энергия «стекает» к реакционным центрам. Спектральные исследования позволили расчленить формы хлорофилла на три основные группы (коротковолновые, длинноволновые и промежуточные) в соответствии с их ролью в поглощении и переносе энергии.
У фотосинтетических бактерий также обнаружены субклеточные частицы, содержащие бактериохлорофилл. Это уплощенные диски диаметром 100 нм, носящие название хроматофоры.
Структуры пигментобелковых комплексов в организации фотосинтетических мембран различных организмов, включая бактерии, водоросли и высшие растения, сходны. Полипептиды хлорофиллобелкового комплекса синтезируются внутри хлоропластов; они состоят из главного полипептида с мол. весом (массой) 73 000 и трех минорных с молекулярным весом (массой) 47 000, 30 000 и 15 000 единиц.
Синтез и обновление пигмента в растущей зеленой ткани протекают с высокой скоростью. С возрастом ткани процесс биосинтеза хлорофилла замедляется. На первых этапах биосинтеза хлорофилла путем конденсации двух молекул δ-аминолевулиновой кислоты формируется порфо-билиноген — производное пиррола, которое в результате ряда превращений дает соединение, содержащее порфириновое ядро — протопорфирин. Из протопорфирина образуется непосредственный предшественник хлорофилла — протохлорофиллид, содержащий атом магния. Затем после присоединения многоатомного спирта фитола образуется хлорофилл.
Этапы от порфобилиногена до протопорфирина и от протопорфирина до хлорофилла а осуществляются по одной из двух схем:

Первая реакция преобладает в листьях этиолированных (то есть выросших в темноте) растений, вторая — в зеленых. Терминальные стадии биосинтеза пигментного аппарата ускоряются при участии единого полиферментного хлорофилл-синтетазного комплекса. В связи с этим естественна зависимость биосинтеза хлорофилла от скорости белкового синтеза и торможения его ингибиторами синтеза белка. Синтез пигментов замедляется также при снижении температуры и полностью прекращается при температуре ниже —2°, тогда как фотосинтез продолжается и при отрицательных температурах, вплоть до —24°. Процесс нарушается при недостаточности железа и избытке марганца.
Образование хлорофилла b происходит последовательно через хлорофилл а путем окисления. Реакция превращения идет на свету; промежуточной стадией является образование фермент-белкового комплекса.
Есть указания на зависимость скорости реакции от работы электронно-транспортной цепи и соотвественно скорости генерации НАДФН и НАДН как доноров водорода. Остаются неясными стадии синтеза на участке включения магния, превращения Mg-порфиринов, а также этерификации фитолом остатка нропионовой кислоты IV пиррольного кольца.
Способность зеленых растений образовывать в процессе фотосинтеза сложные органические вещества из двуокиси углерода и воды определяется присутствием в них хлорофиллов. При этом содержание пигментов хлорофилла а и хлорофилла Ъ не зависит от географических особенностей местности. Содержание хлорофилла а в большей степени подвержеко влиянию физиологических и экологических условий, чем содержание хлорофилла Ъ.
Описаны изменения хлорофиллов в онтогенезе растений. Их содержание возрастает в фазу кущения, в фазу цветения и завязывания плодов. По уровню хлорофилла можно определить готовность растений к цветению. После завершения ростовых процессов накопление хлорофилла прекращается, и обновление молекул пигмента происходит внутри хлоропласта, не будучи связано с образованием новых хлоропластов.
Принцип фотосенсибилизирующего действия хлорофиллов при фотосинтезе был обоснован К. А. Тимирязевым и включает возбуждение пигмента светом с переходом пигмента в синглетное или триплетное состояние и последующими обратимыми фотохимическими изменениями. Хлорофилл на разных этапах может служить фотохимическим донором или акцептором электронов.
Поскольку тетрапиррольным структурам, содержащим комплексно связанный атом железа, принадлежит важная роль в тканевом дыхании млекопитающих (см. Гемоглобин), хлорофилл и его металлопроизводные (т. е. соединения, в структуру которых вместо магния введены медь, железо, цинк, кадмий или серебро) используют в медицине в качестве антигипоксических средств. Металлопроизводные хлорофилла получили название «феофитинаты». Их антигипоксический эффект связывают с тетрапиррольной структурой и присутствием атома металла. Водорастворимые препараты хлорофилла обладают антибактериальной и противовирусной активностью, особенно Ag-феофитинат. Гемопоэтические, общетонизирующие свойства присущи хлорофиллину натрия, который используют также в качестве биостимулятора.
Библиогр.: Годнев Т. Н. Хлорофилл, Его строение и образование в растении, Минск, 1963, библиогр.; Красновский А. А. Уровни светового регулирования фотосинтеза, в кн.: Теоретические основы фотосинтетической продуктивности, под ред. А. А. Ничипорови-ча, с. 23, М., 1972; Мецлер Д. Э. Биохимия, Химические реакции в живой клетке, пер. с англ., т. 1—2, М., 1980; Проблемы биосинтеза хлорофиллов, под ред. А. А. Шлыка, Минск, 1971; Шлык А. А. Метаболизм хлорофилла в зеленом растении, Минск, 1965, библиогр.; Е igenberg К. Ё., С г о a s-m u n W. R. a. Chan S. I. Chlorophyll a in bilayer membranes, Biochim. biophys. Acta, v. 679, p. 353, 1982; Metabolic pathways, ed. by D. M. Greenberg, v. 2, N. Y.— L., 1967; Olson J. M. Chlorophyll organization in green photosynthetic bacteria, Biochim. biophys. Acta, v. 594, p. 33, 1980.
ХЛОРОФИЛЛЫ (от греч. chloros - зеленый и phyllon -лист), прир. макрогетероциклич. пигменты, участвующие в процессе фотосинтеза; относятся к металлопорфиринам (см. Порфирины).
Зеленая окраска растений обусловлена присутствием хлорофиллов, локализованных во внутриклеточных органеллах (хлоропластах или хроматофорах) в виде пептидных комплексов.
Формально хлорофиллы представляют собой производные порфина, молекулы к-рых содержат циклопентаноновое кольцо, конденсированное с порфириновым макроциклом, центральный атом Mg и разл. заместители; одно или два пиррольных цикла в молекулах частично гидрированы, см., напр., ф-лу I. В пиррольном кольце D молекул хлорофиллов к остатку пропионовой к-ты обычно присоединены остатки высокомол. изопреноидных спиртов, к-рые придают хлорофиллам способность встраиваться в липидные слои мембран хлоропластов. Для хлорофиллов, как и для порфиринов, используется номенклатура ИЮПАК или Фишера.
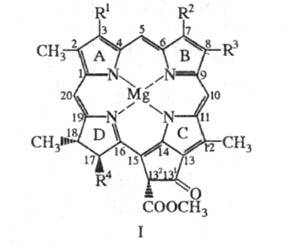
Хлорофилл b: R 1 = СН = СН2, R 2 = СНО, R 3 = C2H5, R 4 = CH2CH2C(O)Y
Из бурых и диатомовых водорослей выделены хлорофиллы а и с, из красных морских водорослей - хлорофиллы а и d.
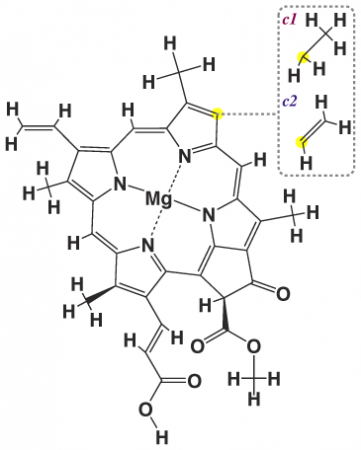
Х лорофиллы группы с (c1, с2 и c3, ф-ла II) в отличие от др. хлорофиллов содержат негидрированный порфириновый макроцикл и остаток неэтерифицированной акриловой к-ты. Находясь в морских водорослях в виде белковых комплексов, хлорофиллы этой группы выполняют в фотосинтезе роль светособирающих антенн.
В большинстве фотосинтезирующих бактерий обнаружены бактериохлорофиллы (БХ), отличающиеся от хлорофилла а типом макроцикла и замещающими группами в цикле. Они имеют несколько модификаций: так, из пурпурных бактерий выделены БХ а и b, из зеленых бактерий - БХ а, с, d и е, из серных бактерий - БХ с, d и е; обнаружены также фотосинтезирующие бактерии, содержащие БХ g.
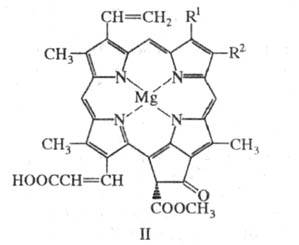
Хлорофилл c1: R l = CH3, R 2 = C2H5 Хлорофилл с2: R 1 = CH3, R 2 = CH = CH2 Хлорофилл с3: R 1 = СООСН3, R 2 =CH=CH2
В основе БХ a, b и g (т. наз. собственно БХ; ф-ла III) лежит тетрагидропорфириновый макроцикл, содержащий в качестве эфирных групп (Y) остатки фитола, геранилгераниола (СН3)2С = СН(СН2)2С(СН3) = СН(СН2)2С(СН3) = СН(СН2)2С(СН3) = СНСН2ОН и 2,10-фитадиенола (СНO2СН(СН2)3С(СН3) = СН(СН2)2СН(СН3)(СН2)3С(СН3) = СНСН2ОН - для БХ а и b; БХ g содержит остатки фарнезола
(СН3)2С = СН(СН2)2С(СН3) = СН(СН2)2С(СН3) = СНСН2ОН и геранилгераниола. При выделении из ацетона или метанола (особенно в присут. оснований) БХ а и b эпимеризуются по атому С-13 2 с образованием эпимеров БХ а' и b'.
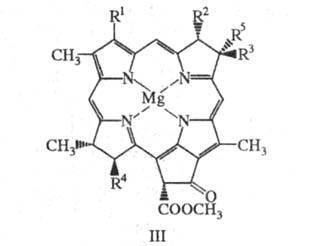
Бактериохлорофилл a: R 1 = СОСН3, R 2 = СН3, R 3 = С2Н5, R 4 = CH2CH2C(0)Y, R 5 = Н
Бактериохлорофилл b: R 1 = СОСН3, R 2 = СН3, R 3 + R 5 = (=СНСН3), R 4 = CH2CH2C(O)Y
Бактериохлорофилл g: R 1 = СН = СН2, R 2 = CH3, R 3 + R 5 = (= CHCH3), R 4 = CH2CH2C(O)Y
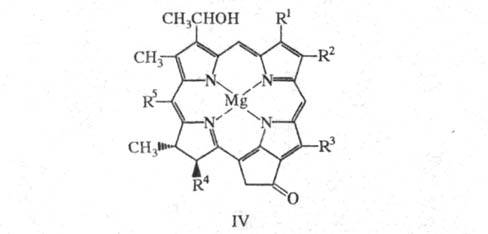
Для БХ с, d и е (ф-ла IV), первоначально называемых хлоробиум-хлорофиллами, характерно наличие дигидропорфиринового макроцикла, a -гидроксиэтильной группы в положении 3 и разл. алкильных (от С1 до С5) заместителей в положении 8; эфирные группы (Y) - остатки 2,6-фитадиенола (СН3)2СН(СН2)3СН(СН3)(СН2)3С(СН3) = СН(СН2)2С(СН3) = СНСН2ОН и 2,16,20-фитатриенола (CH3)2C = СН(СН2)2С(CH3) = СН(СН2)2СН(СH3)(СН2)3 -С(СН3) = СНСН2ОН.Х лорофиллы- высокоплавкие интенсивно окрашенные кристаллы от зеленого до темно-красного и черного цветов; т. пл. хлорофилла а 117-121 °С, хлорофилла b - 124-125 °С; т. разл. многих хлорофиллов более 300 °С. Хлорофиллы хорошо раств. гл. обр. в полярных орг. р-рителях (ДМСО, ДМФА, ацетон, спирты, диэтиловый эфир), плохо -в петролейном эфире, не раств. в воде. В УФ спектрах для многих хлорофиллов характерно наличие
НЕКОТОРЫЕ ХАРАКТЕРИСТИКИ ХЛОРОФИЛЛОВ И БАКТЕРИОХЛОРОФИЛЛОВ
Электронный спектр, x 10 -3 или отношение интенсивностей к основному пику)
Строение хлорофилла
Фотосинтез у высших зеленых растений протекает в специализированных внутриклеточных органеллах — хлоропластах, способных доставлять энергию для метаболических процессов только в дневные часы.
Среди светособирающих первостепенная роль принадлежит двум молекулярным формам хлорофилла – а и b (рис. 34).
Рис. 34. Строение хлорофиллов a и b
В молекуле хлорофилла, присутствующего в хлоропластах клеток зеленых растений, содержится четыре пирольных кольца (I—IV), одно из которых (IV) находится в восстановленной форме. Имеется еще и V непирольное кольцо. Длинная изопреноидная боковая цепь в молекуле хлорофилла представляет собой остаток спирта фитола С20Н39ОН — производного ненасыщенного углеводорода изопрена, который присоединен сложноэфирной связью к карбоксильной группе заместителя в кольце IV. Четыре центральных атома азота в молекуле хлорофилла координационно связаны с ионом Мg 2+ двумя основными и двумя наведенными дополнительными валентностями.
По своей химической природе хлорофилл представляет собой сложный эфир двухосновной кислоты и двух спиртов — метилового и фитола. Остаток фитола придает молекуле хлорофилла липидные свойства и обеспечивает закрепление и ориентацию его в природной тилакоидной мембране хлоропласта.
Хлорофилл при гидролизе омыляется с образованием щелочных солей хлорофилла и спиртов. При действии слабых кислот ион магния вытесняется из центра порфиринового ядра и замещается водородом. В результате образуется соединение бурого цвета, которое называется феофитином.
В фотосинтезирующих клетках высших растений всегда присутствуют хлорофиллы нескольких типов, важнейшие из которых хлорофиллы а (С55Н72О5N4Mg) и b (С55Н70О6N4Мg), отличающиеся только тем, что у хлорофилла b вместо метильной в кольце II содержится формильная группа (О=С—Н). Хотя хлорофиллы а и b окрашены в зеленый цвет, их спектры поглощения несколько различаются. У большей части высших зеленых растений количество хлорофилла а примерно вдвое превышает количество хлорофилла b.
Относительное содержание хлорофиллов и каротиноидов у разных видов растений заметно варьирует. Именно от их соотношения зависит характерная окраска фотосинтезирующих клеток. Содержание хлорофилла на сухой вес растения составляет примерно 1%.
Система из пяти колец, образующая кольцо больших размеров вокруг атома Мg 2+ , придает молекуле способность поглощать свет. Атом магния собирает молекулы хлорофилла в ассоциации, что облегчает более полное улавливание света.
В тилакоидных мембранах хлоропластов кроме хлорофилла присутствуют также и вспомогательные светопоглощающие пигменты – каротиноиды, окрашенные в желтый, оранжевый или красный цвет. Среди них наиболее важные – красный пигмент b-каротин (провитамин А) и желтый пигмент ксантофилл, или лютеин. Он является главным ксантофиллом листьев.
Каротиноиды поглощают свет в ином диапазоне, нежели хлорофиллы, поэтому они функционируют как световые рецепторы, дополняющие хлорофиллы.
Читайте также: